随着肥胖发病率的增加,使用小鼠作为研究饮食诱导肥胖(DIO)模型的人数显著增加。虽然许多小鼠品系对DIO敏感,但不同品系之间的敏感性差异很大。例如,SWR/J和CAST/Ei倾向于具有DIO抗性,而C57BL/6品系则高度敏感。因此,在肥胖研究中,C57BL/6J小鼠经常使用高脂饮食(HFD)饲喂用于建立肥胖模型。
许多因素会影响 C57BL/6 小鼠的体重,其中影响最大和最众所周知的因素是年龄、性别和饮食。以前对 C57BL/6 小鼠的 HFD 诱发肥胖的研究使用了相对较小的样本量或较短的研究持续时间,仅报告了雄性的结果或使用了非常高的脂肪成分(大约 60% 的卡路里来自脂肪)。增加样本量有助于了解数据间的差异,即使在已报告的相对容易肥胖造模的品系中也是如此。有人提出,雄性C57BL / 6J小鼠在短期HFD喂养后,在稳定的肥胖表型方面存在很大差异,尽管它们是近交系且据称是等基因遗传状态的,但很少有研究报告长期结果或雌性 C57BL/6 小鼠对 DIO 的反应 。此外,60% 的热量来自脂肪的饮食比脂肪含量较低的饮食(如 45%)更有效地诱导小鼠肥胖;然而,60% 的热量来自脂肪远高于人类正常膳食脂肪摄入量,特别是当它主要来自单一脂肪来源时 。 NHANES 膳食营养素摄入量调查报告称,美国成年男性和女性能量摄入的 30-40% 来自脂肪。除了这些因素之外,产仔数和断奶体重与 C57BL/6J 小鼠的肥胖呈弱正相关。
在本文中,我们使用了较大的样本量,来研究雄性和雌性 C57BL/6J 小鼠在长期 HFD 喂养后体重、食物摄入量和身体成分的变化。
方法
动物
六周龄 C57BL/6J 小鼠(277只雄性和 278只雌性),从杰克逊实验室获得,并以5只/笼的密度使用标准饮食饲养一周( Teklad Global 16% 蛋白质啮齿动物饮食)。随后将小鼠单独饲养一周,然后从 8 周龄开始喂食 HFD(D12451,45% 千卡脂肪和 20% 蛋白质。将 一个月前获得的另一组 30 只相同年龄和品系的小鼠(15只雄性和15只雌性)作为低脂肪对照组。 在适应一周后,这 30 只小鼠被单独饲养,随后从 8 周龄开始喂食低脂饮食(LFD;D12450B,10% 大卡脂肪和 20% 大卡蛋白质)。
所有小鼠都饲养在带有通过过滤系统的标准小鼠笼中,笼中装有 200 g 灭菌的灭菌刨花垫料和和压缩纸卷垫料。饲养室是一个无特定病原体的动物设施,有哨兵监测。 所有的笼子都保持在通风架系统上。笼具中的垫料和水,每月更换一次,笼子在架子上的位置每月更换一次,以减少环境的影响(光照强度、噪音、振动等)。 从早上 6 点开始,动物房保持 12:12 小时的明暗循环,环境温度为 22±1℃,湿度为 50%。 所有程序均按照阿拉巴马大学伯明翰分校的机构动物护理和使用委员会 (IACUC) 执行。
措施
第一个月每周测量体重,此后每四周测量一次体重,直到 43 周龄(除非特殊指定,所有周数表示小鼠年龄)。 每两周更换一次食物,每两周测量一次。 食物摄入量是通过从称重的等分试样中减去剩余的食物(包括笼中任何溢出的食物)两周时间来获得的。 根据 HFD 的 4.73 kcal/g 和 LFD 的 3.85 kcal/g(热量值从研究饮食获得)计算能量摄入。 如所述,在 8、23 和 43 周龄时使用定量磁共振 (QMR) 在体内测定身体成分(脂肪和无脂肪质量)。 体重变化的热量含量计算为无脂肪组织为 1.8 kcal/g,脂肪组织为 8 kcal/g。
根据大学和 IACUC 的政策,对濒死的小鼠,包括患有溃疡性皮炎或类似皮肤损伤的小鼠 (n=29) 或无法进食/饮水的小鼠(一只雌性)实施安乐死。 一只雄性小鼠在研究过程中自发死亡。 在 10 个月大之前,共有 23 只雌性和 8 只雄性小鼠死亡或被移走; 因此,554 只小鼠被纳入数据分析(255 只 HFD 雌性、269 只 HFD 雄性、15 只 LFD 雌性和 15 只 LFD 雄性)。
统计分析
所有统计分析均使用 SAS(9.3 版;SAS Institute,Cary,NC)进行。 学生 t 检验或重复测量方差分析和 Tukey-Kramer 调整用于多重比较,在适当的情况下用于确定体重、食物摄入量和身体成分随时间的显着差异。 重复测量协方差分析(ANCOVA)用于比较组间体重的变化。 为了比较体重和体重增加,我们采用三次多项式模型。 时间(周)是一个连续变量,有平方项和立方项,模型包括饮食和性的主要影响,以及它们的双向交互作用和它们与时间的交互作用。 这允许每种饮食和性别组合具有明显的三次生长曲线,并允许在两种饮食不常见的时间点进行比较。 重复测量使用空间功率协方差结构建模,因为测量是在不相等的时间间隔进行的。
采用非参数Mann-Whitney检验对不同性别的人体成分进行比较。皮尔逊相关被用来测试体重、能量摄入和身体成分之间的相互关系。多变量方差分析用于预测每种性别、体重和脂肪质量随时间的变化,体重(或脂肪质量)作为因变量,三个基线特征作为自变量。统计显著性标准为P<0.05(双尾),使用经多重比较调整的P值。
结果
体重
从基线(8 周龄)开始,HFD 和 LFD 小鼠随着时间的推移显着增加体重(all P < 0.0001)(图 1A),具有显着的饮食和性别影响。 从第 8 周到第 43 周,HFD 男性的平均体重增加为 20.53 克,HFD 女性为 12.55 克,而 LFD 男性为 10.5 克,LFD 女性为 5.62 克。 在每种性别中,HFD 雄性小鼠从第 14 周开始明显比 LFD 小鼠重(P = 0.0221),而 HFD 雌性小鼠在第 27 周时与 LFD 雌性小鼠表现出统计学差异(P = 0.0076)。
至于体重的增加(图 1B),HFD 喂养的雄性和雌性之间的显着差异在第 10 周首次明显(P = 0.0027)。 从第 14 周开始,HFD 雄性的体重增加仍然显着大于 LFD 雄性和两个磁性组(第 14 周 HFD 男性和 LFD 男性之间的 P = 0.0028;所有其他 P < 0.0001)。 以类似于同性别内的绝对体重比较的方式,HFD 男性的体重变化从第 14 周开始与 LFD 男性有显着差异(P = 0.0028),之后仍然存在显着差异(所有 P < 0.0001 ),而女性从第 23 周开始(P = 0.0178)并保持显着差异(第 27 周 P = 0.0009,其余周 P < 0.0001)。 与 LFD 男性和 HFD 女性之间绝对体重的显着差异(从第 8 周到第 39 周)相比,这两组在所有周内的体重增加在统计学上相似。
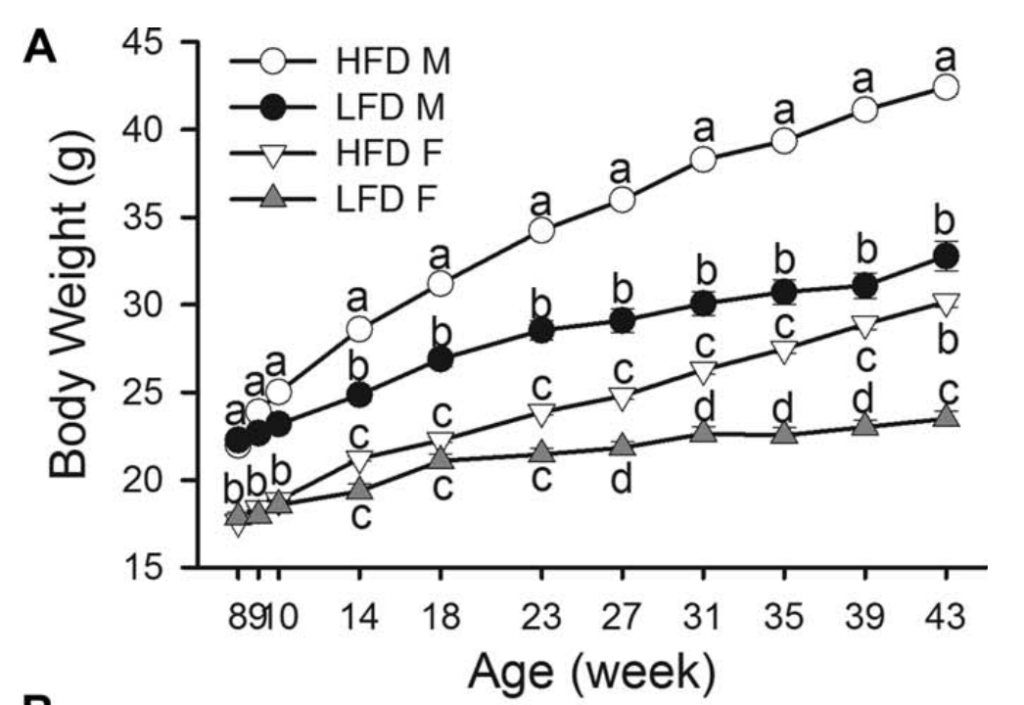
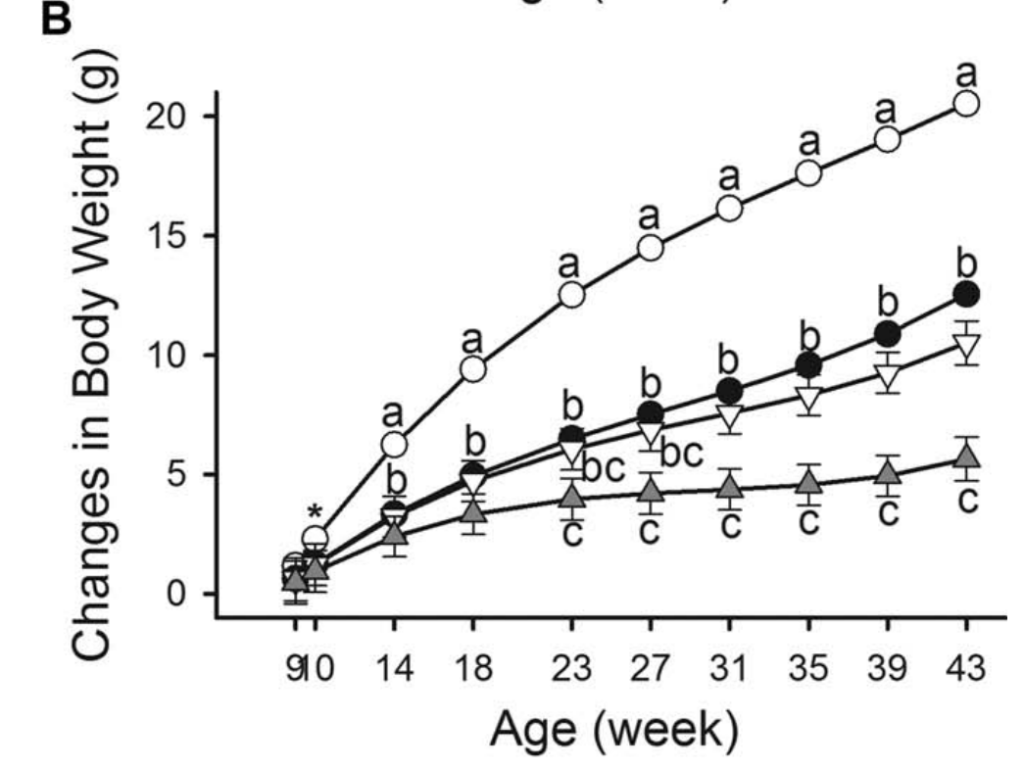
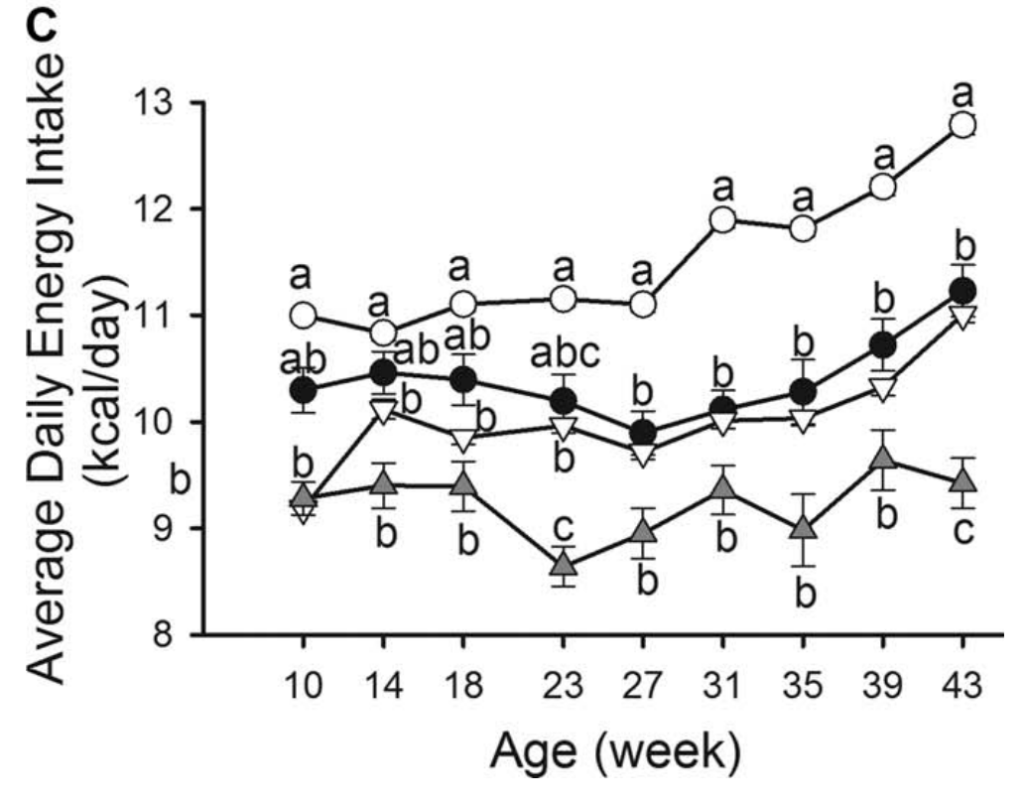
图1:体重和能量摄入量。(A)体重随时间变化。高脂肪饮食的雄性小鼠(HFDM;45%千卡脂肪;n=269)、低脂饮食的雄性小鼠(LFDM;10%千卡脂肪;n=15)、高脂饮食的雌性小鼠(HFDF;n=255)和低脂饮食的雌性小鼠(LFDF;n=15)。(B)所有组的体重变化(每周的体重减去基线体重)。(C)各组患者的平均每日能量摄入量随时间而变化。HFD男性和女性第18周的体重和能量摄入量为第17周和第19周的平均值,使其与LFD相当。*表示在第10周时,HFD女性和HFD男性之间存在显著差异。每个时间点的不同字母表示在P<为0.05处有显著差异。误差条表示SE。
能量摄入量
在 HFD 中,男性的能量摄入明显高于女性(所有周 P < 0.0001); 然而,在 LFD 中,直到第 43 周才有显着差异(P=0.0103)。 HFD 雄性和雌性小鼠能量摄入的最终测量值显着高于初始测量值(均 P < 0.0001),但 LFD 小鼠没有显着差异(雄性 P = 0.9494,雌性 P = 1.0000)。 HFD 和 LFD 动物之间每日能量摄入的显着差异首先在第 27 周的雄性中得到证实(P=0.0391),但在第 23 周(P=0.0091)和第 43 周(P=0.0002)再次出现在雌性中(P=0.0002)。 图 1C)。
整个研究的平均每日能量摄入与所有组的总体重增加显着相关(图 2A 和 C),并且与增加的体重(脂肪质量+无脂肪质量)的热量含量类似地相关(图 2B 和 D)。 HFD 女性的平均每日能量摄入与总体重增加之间的相关性(R=0.7268,P < 0.0001)强于 HFD 男性(R=0.3748,P < 0.0001)。 同样,平均每日能量摄入量与体重增加的热量含量之间存在显着相关性(HFD 女性的 R = 0.6989,P < 0.0001;HFD 男性的 R = 0.3377,P < 0.0001)。
身体构成
除了 LFD 女性的脂肪量(P=0.6597)(图 3A 和 B)外,脂肪量和无脂肪量均随时间显着增加(所有 P < 0.0001)。在基线时,无论是 HFD (P=1.0000) 还是 LFD (P=1.0000),男性和女性的脂肪量都没有统计学差异。然而,对于 HFD (P < 0.0001) 和 LFD (P < 0.0001),男性和女性的无脂肪质量存在显着差异。在第 23 周和第 43 周,雄性比雌性具有更大的无脂肪质量,并且 HFD 小鼠比 LFD 小鼠具有更大的脂肪量。对于第 8 周至第 23 周的体重变化,在 HFD(10.15 g 对 3.14 g,P < 0.0001)和 LFD(4.36 g 对 0.98 g,P =0.0066);在 HFD (P=0.9030) 或 LFD (Tukey-Kramer 调整后 P=1.000,未调整 P=0.9734) 中,男性和女性在无脂肪质量变化方面没有显着差异。对于第 23 周至第 43 周的体重增加,HFD 男性和女性(P=0.9892)或 LFD 男性和女性之间(P=0.1986)的脂肪量增加没有显着差异; HFD 男性和女性之间的无脂肪质量变化有显着差异(1.95 g 对 1.26 g,P < 0.0001),但 LFD 男性和女性之间没有显着差异(P = 0.8005)。
虽然无脂质量的数量随着时间的推移而显著增加,但四组的无脂质量的变化都保持相对稳定。这可以从变异系数中得到证明,从第43周LFD男性的0.042到第8周HFD女性的0.070。相比之下,脂肪质量的变异性随着时间的推移而增加,在第8周时变异系数最低,为0.138,在第43周时最高,为0.390。这一点如图3C-E所示。所有组的脂肪质量和无脂肪质量在8、23和43的分布如图4所示。如图4E和F,男性和女性的脂肪质量显著增加的变化(P<0.0001),但无脂肪质量的方差没有显著变化,除了HFD男性从23周到43周(P<0.0001)。对于LFD小鼠,脂肪质量和无脂肪质量都没有随着时间的推移而增加,除了LFD雄性的脂肪质量差异,从第8周到第23周显著增加(P<0.0001)。
支持信息 图 2 说明了从基线(第 8 周)到研究结束(第 43 周)体重增加与脂肪量和无脂肪量增加之间的关系。 对于 HFD 男性 (R=0.9692, P < 0.0001)、LFD 男性 (R=0.9349, P < 0.0001) 和 HFD 女性 (R = 0.9791,P < 0.0001)但不适用于 LFD 女性(R = 0.3901,P = 0.1506)。 同样,但在较小程度上,无脂肪质量增加(D 无脂肪质量)也与 HFD 男性(R = 0.7456,P < 0.0001)、HFD 女性(R = 0.5294,P < 0.0001)的 D 体重显着相关 ),和 LFD 女性 (R=0.6822, P=0.0051) 但不是 LFD 男性 (R=0.5074, P=0.0535)。 此外,在大多数时间点,所有四组的绝对脂肪量和无脂肪量之间存在显着相关性(支持信息表 1)。

图2能量摄入与总体重增加或体重增加的热量含量的相关性。(A)HFD男性和LFD男性的平均每日能量摄入和总体重增加和体重增加的(B)热量含量;(C)HFD女性和LFD女性的平均每日能量摄入和总体重增加以及体重增加的(D)热量含量
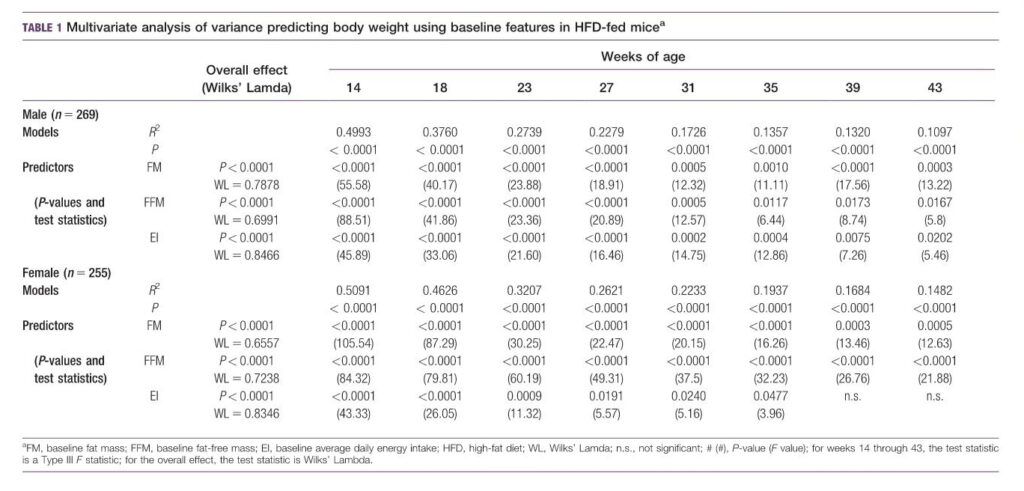
对HFD小鼠体重(增重)预测的多变量方差分析
考虑到在研究过程中观察到的最终体重(和脂肪质量)的巨大变异性,我们将脂肪质量、无脂肪质量和平均每日能量摄入的基线值作为DIO反应的预测因素(或解释变量)。体重预测的多变量方差分析(表1)表明,这些基线测量对预测未来体重具有显著性,但其显著性具有性别特异性。
在HFD雄性小鼠未来体重的这三个预测因子中,它们在整个研究过程中都是未来体重的显著阳性预测因子(所有总体效应P<0.0001)。对于HFD雌性小鼠,基线脂肪质量和无脂肪质量是所有时间点的显著预测因子(总体效应P<0.0001);基线能量摄入在第35周是一个显著的预测因子(在第35周是P5 0.0477 )。
对于脂肪质量的预测(表2),基线脂肪质量对两性都是一个显著的预测因子(总体效应P<0.0001)。基线无脂肪对HFD女性的两种未来测量都是一个重要的预测因素,但仅对HFD男性在第23周的测量具有显著性。基线能量摄入仅在HFD男性第23周(P<0.0001)和HFD女性(P5 0.0054)是一个显著的预测因子。
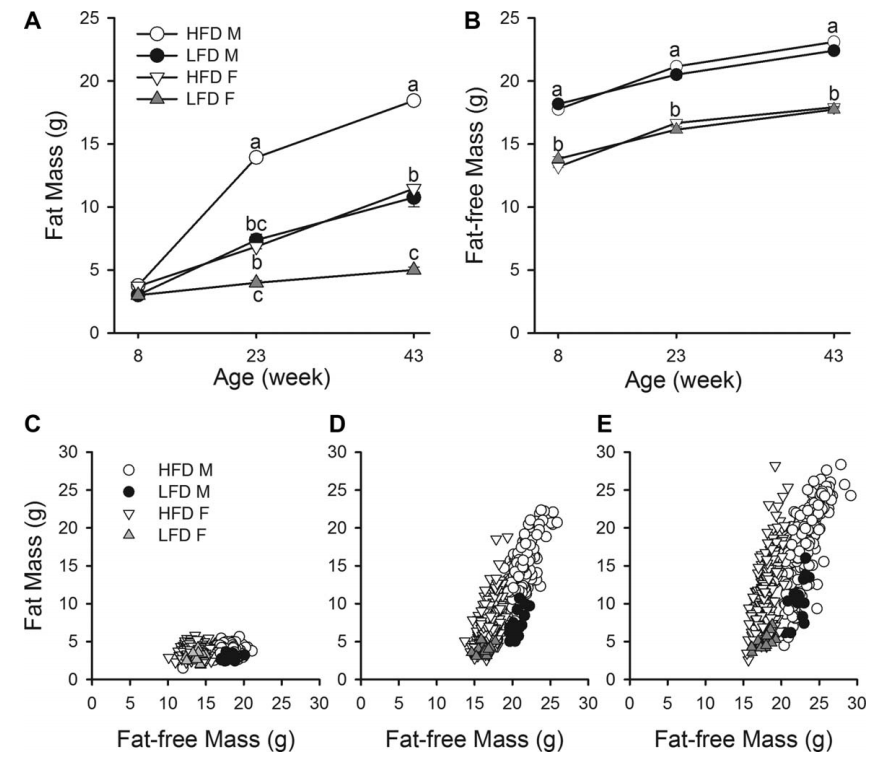
图3:身体成分的变化。(A)所有组的脂肪质量(平均SE);所有组的(B)无脂肪质量(平均SE);在(C)第8周、(D)第23周和(E)第43周时,所有组的身体组成(脂肪质量与无脂肪质量)分散。不同字母表示P<0.05有显著差异。
讨论
本研究中提供的数据来自一项正在进行的长寿研究的DIO阶段,该研究使用的是有肥胖倾向的C57BL/B6J小鼠。所用饮食中的脂肪含量与美国人口(12,13)中的脂肪摄入量相似。虽然大多数已发表的关于C57BL/6小鼠中DIO的文献只关注雄性,但我们使用了两性。与其他研究(6,16)一致,我们的结果显示,雄性C57BL/6J小鼠对DIO敏感,而雌性小鼠的敏感程度较轻,且速度较慢。此外,在体重的变化和分布、能量转换为体重和身体组成的变化以及具有基线特征的体重预测等方面也存在性别差异。

图4 第8周、第23周、第43周的体成分分布。直方图分布(用30个箱子绘制,脂肪质量为0到30g,无脂肪质量为10到30g)。(A)HFD男性脂肪质量;HFD女性(B)脂肪质量;HFD男性(C)无脂肪质量;HFD女性(D)无脂肪质量。(E)所有组的脂肪质量和(F)所有组的无脂肪质量的个体散点图分布。
雄性 C57BL/6 小鼠一直是用于 DIO 研究的标准动物 (1-4),但很少有研究报告雌性 C57BL/6 对高脂肪 DIO 的反应,其局限性包括样本量小或研究持续时间短。 Tortoriello 等人 (17,18) 证明在 C57BL/6J 品系中存在明显的雌性对 DIO 的性别特异性抗性,其中 24% kcal 脂肪饮食持续 20 周 (n=20) 和 35% 脂肪饮食持续 24 周 (n=10) 和推测延长喂养或高脂肪饮食可能更有效。后来的一项研究 (11) 发现,饮食中脂肪含量越高,体重越高,我们观察到45%大卡脂肪饮食12周(n=50)体重增加24%,脂肪含量更高(60%)时体重增加39%,该研究不支持性别特异性肥胖抵抗的概念。我们发现,在高脂喂养 35 周后,我们的 HFD雌性小鼠(n = 255) 体重平均增加了 71%,而雄性小鼠 (n = 269) 增加了 93%。在最终体重分布中,雄性偏左(偏度=20.284),而雌性偏右(偏度= 0.659)。这表明小部分雌性小鼠普遍倾向于大大超过其预期体重增加,而一小部分雄性小鼠未达到预期体重增加。当雄性和雌性小鼠喂食 HFD 和 LFD 时,雄性小鼠比雌性更早观察到两种饮食之间的显着差异,这表明雄性对 HFD 的反应比雌性更快。其他饮食相同或相似的人 (6,19) 也观察到男性 HFD 和 LFD 之间体重的早期差异。雌激素水平 (20)、性别特异性瘦素抵抗 (4,21) 和总体运动活动的差异 (16) 可能导致对 DIO 的反应存在性别差异,但在本研究中未对它们进行测试。
我们发现,在长期 HFD 喂养后,动物的体重和脂肪量差异很大,这与其他研究的结果一致 (8,9,11,22)。随着小鼠变大,雄性和雌性的脂肪量变化大于无脂量的变化,这表明无脂量受饮食控制的影响比脂肪量小。确定体重或脂肪量变化何时发生增加以及是什么导致这种遗传上相同的近交系小鼠品系的变化非常重要。科扎等人 (9) 表明,即使在 LFD 喂养数周后,体重的变化也发生在幼年的时候,并且这种变化在热量限制的情况下仍然存在。我们通过分别分析在所有同性 HFD 动物的前 10% 或后 10% 中完成研究的 HFD 动物来检验这一现象。在基线时,雄性小鼠 (P < 0.0001) 和雌性小鼠 (P < 0.0001) 之间存在显着差异(支持信息图 1A 和 B),这表明在接受 HFD 之前已经存在体重变化。此外,即使在 HFD 喂养下,底部 10% HFD 小鼠(雄性和雌性)的体重在大多数时间点与 LFD 小鼠的体重没有显着差异。
有人认为,食物摄入有助于小鼠肥胖的长期变化。在我们的数据中,平均每日能量摄入与总体重增加(或体重增加的热量含量)之间的显著相关性支持了这一点。有趣的是,能量摄入和体重增加(或体重增加的热量含量)之间的相关性在HFD雌性小鼠中比在HFD雄性小鼠中更强。此外,平均每日能量摄入的前10%HFD动物显著高于底部10%的研究雄性小鼠(所有P<0.0001)和14周雌性小鼠(10周P=0.0798 ,所有其他P<0.0001)(支持信息图1C和D)。这表明初始体重较低的小鼠和体重较大的小鼠对HFD没有反应。
鉴于此 DIO 模型的可变性,如果为未来体重或体脂预测建立了参数,这将是有用且具有成本效益的。张等人 (8) 研究了预测 60 只 C57BL/6J 小鼠体重增加的非遗传变异性的潜在因素,发现 HFD 诱导的肥胖与基线脂肪量、无脂肪量和身体活动有关。他们使用年龄相近的雄性C57BL/6J 小鼠接受相同的 HFD 16 周,发现初始体脂是体重增加变异性的最强预测因子,与 HFD 喂养时间无关,基线身体活动和无脂肪在 16 周的研究中,体重与后期体重增加有关。然而,基线食物摄入量、静息代谢率和体温在任何时间点都不是显着的预测因子。我们还回顾性地研究了基于三个基线特征的体重预测,包括能量摄入、脂肪量和无脂肪量。我们发现这些特征是未来两性体重的重要预测指标,但基线能量摄入对女性仅部分显着。此外,基线脂肪量是男女未来体脂的重要预测指标,而基线无脂肪量仅对整个研究中的女性显着。这些结果表明生长发育的性别差异。总之,雄性和雌性 C57BL/6J 小鼠在生长模式上存在差异,除了雄性具有更高的体重、脂肪量和无脂肪量。男性和女性的脂肪量差异很大,并且这种差异随着时间的推移而增加,而无脂肪量的变化保持相对稳定。多元方差分析模型表明,脂肪量、无脂肪量和能量摄入等基线特征可以作为未来体重估计的预测因子。研究人员应考虑这种 DIO 易感小鼠品系的变异和性别差异。在近交系小鼠品系中寻找 HFD 引起的肥胖个体差异的原因的研究是必要的。

原文下载:
[download id=”443″]
shuyushengwu
请到「后台-用户-个人资料」中填写个人说明。






© 2025. All Rights Reserved. 沪ICP备2020028303号-1  沪公网安备 31011002005293号
沪公网安备 31011002005293号


